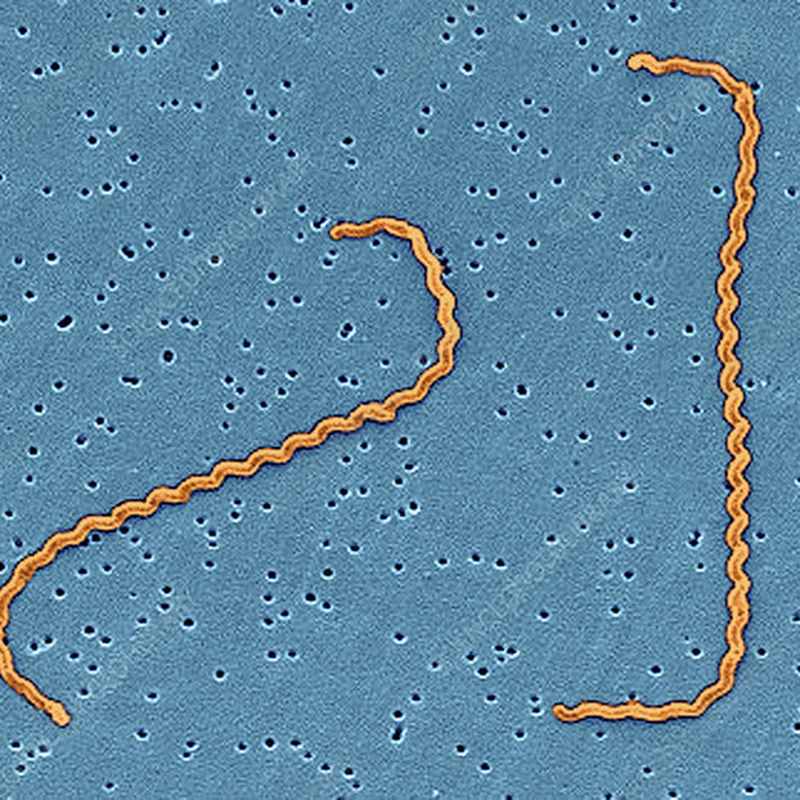
Leptospira (combinazione della parola greca λεπτος, leptos, "sottile", e della parola latina spira, con lo stesso significato dell'italiano "spira") è un genere di batteri appartenente al phylum Spirochaetes. Ha forma di filamento elicoidale ad estremità uncinate e possiede 2 flagelli che lo rendono mobile. Alcune specie sono patogeniche (v. leptospirosi), altre sono semplicemente saprofitiche.
Leptospira fu osservato per la prima volta nel 1907 nei tessuti renali di un paziente morto per quella che fu descritta erroneamente come febbre gialla.
Tassonomia
Leptospira, insieme ai generi Leptonema e Turneria, appartiene alla famiglia delle Leptospiraceae.
Il genere è diviso in 20 specie in base a studi sul DNA come segue:
Specie patogeniche:
- Leptospira interrogans
- Leptospira kirschneri
- Leptospira noguchii
- Leptospira alexanderi
- Leptospira weilii
- Leptospira genomospecies 1
- Leptospira borgpetersenii
- Leptospira santarosai
- Leptospira kmetyi
Specie intermedie o opportunistiche:
- Leptospira inadai
- Leptospira fainei
- Leptospira broomii
- Leptospira licerasiae
- Leptospira wolffii
Specie non patogeniche:
- Leptospira biflexa
- Leptospira meyeri
- Leptospira wolbachii
- Leptospira genomospecies 3
- Leptospira genomospecies 4
- Leptospira genomospecies 5
Il genere viene anche diviso in sierotipi in base all'affinità antigenica. Attualmente vengono riconosciuti oltre 200 sierotipi; qualche sierotipo è trasversale a due o più specie.
Morfologia
Nonostante l'alto numero di sierotipi, tutti i membri del genere hanno morfologia simile: sono batteri a forma di spirale, lunghi 6-20 µm, con un diametro di 0.1 µm (distanza tra le spire di circa 0.5 µm). Una o entrambe le estremità sono di solito uncinate.
Poiché sono così sottili, esemplari di Leptospira viventi sono osservati meglio mediante microscopio in campo oscuro.
Struttura cellulare
Leptospira possiede un rivestimento simile alla parete cellulare dei Gram-negativi, formato da una membrana citoplasmatica, uno strato di peptitoglicano e una membrana esterna. Tuttavia, lo strato di peptidoglicano è associato con la membrana citoplasmatica piuttosto che con quella esterna, un arrangiamento che è esclusivo delle spirochete. I due flagelli di Leptospira, emergenti dalle estremità del batterio, si estendono all'interno dello spazio periplasmico, e sono indispensabili per la motilità della Leptospira.
La membrana esterna contiene diverse lipoproteine e proteine transmembrana. Come previsto, la composizione proteica della membrana esterna differisce quando si confrontano ceppi di Leptospira cresciuti in un mezzo artificiale con ceppi di Leptospira isolati da un animale infetto. Diverse proteine della membrana esterna della Leptospira hanno mostrato la proprietà di legarsi alla matrice extracellulare dell'ospite e al fattore H. Queste proteine potrebbero giocare un ruolo importante nell'adesione della Leptospira ai tessuti dell'ospite e nella resistenza al complemento, rispettivamente.
La membrana esterna della Leptospira, analogamente a quella dei batteri Gram-negativi, contiene lipopolisaccaridi (LPS). Differenze nella struttura altamente immunogenica degli LPS spiegano la presenza di numerosi sierotipi di Leptospira.. Di conseguenza, l'immunità è sierotipo specifica; i vaccini attuali per la leptospirosi, che consistono di uno o più sierotipi di Leptospira endemici nella popolazione, proteggono solo contro i sierotipi contenuti nella preparazione. Il lipopolisaccaride (LPS) leptospirale ha mostrato una bassa attività come endotossina.. Un aspetto insolito dei lipopolosaccaridi leptospirali è che attivano le cellule ospiti via TLR-2 piuttosto che TLR4. La struttura unica del lipide A nella molecola di LPS può spiegare queste osservazioni. Infine, l'antigene O della molecola LPS di L. interrogans differisce in un animale con infezione acuta rispetto a un altro con infezione cronica. In ogni caso, non è ancora noto, se esiste, il ruolo che questi cambiamenti dell'antigene O assumono nell'instaurarsi o nel mantenimento di infezioni acute o croniche.
Habitat
Esemplari di Leptospira sono stati trovati in ambienti diversissimi, in tutto il mondo eccetto l'Antartide.
La presenza di acqua o di forte umidità e il pH neutro (6.9-7.4) sono essenziali per la sopravvivenza di questi batteri. L'habitat naturale usuale è costituito da acque stagnanti.
Nutrizione
Leptospira è coltivata a 30 °C sul terreno di Ellinghausen-McCullough-Johnson-Harris (EMJH), che può essere arricchito con siero di coniglio 0,2-1% per aumentare la crescita di ceppi esigenti. La crescita di Leptospire patogene in un terreno artificiale come l'EMJH si evidenzia in 4-7 giorni, mentre la crescita di ceppi saprofitici avviene in 2-3 giorni. Per le specie patogene, la temperatura minima di crescita è di 13-15 °C. Poiché la temperatura minima di crescita delle specie saprofite è di 5-10 °C, questa differenza può essere usata per distinguere le specie saprofitiche di Leptospira da quelle patogene. Il pH ottimale di crescita di Leptospira è 7.2-7.6.
Le Leptospire sono organismi aerobi nei quali la principale sorgente di carbonio ed energia durante la crescita in vitro è rappresentata da acidi grassi a catena lunga, che sono metabolizzati mediante beta-ossidazione. Gli acidi grassi sono presenti nel terreno EMJH sotto forma di Tween. Nel suddetto terreno gli acidi grassi sono legati dall'albumina e rilasciati lentamente nel mezzo per prevenire il loro accumulo tossico.
Come la maggior parte dei batteri, Leptospira richiede ferro per la crescita. L. interrogans e L. biflexa hanno la capacità di acquisire il ferro in differenti forme. L. biflexa possiede un recettore TonB-dipendente richiesto per l'utilizzazione della forma ferrosa del ferro, ed un gene ortologo del recettore è stato identificato anche nel genoma di L. interrogans. L. interrogans può anche ricavare il ferro dall'eme, che è legato alla maggior parte del ferro presente nel corpo umano. Sulla superficie di L. interrogans è stata identificata la proteina HbpA emina-legante, che può essere coinvolta nella cattura dell'emina. In alcune specie patogene (L. biflexa) non è presente la proteina HbpA, ma un'altra proteina emina-legante, la LipL41, che può spiegare la loro capacità di usare l'emina come sorgente di ferro. Sebbene non secernano siderofori, L. biflexa and L. interrogans sono capaci di ricavare il ferro dai siderofori secreti da altri microrganismi.
Note
Voci correlate
- Leptospirosi
Altri progetti
- Wikimedia Commons contiene immagini o altri file su Leptospira
- Wikispecies contiene informazioni su Leptospira
Collegamenti esterni
- (EN) Kenyon College MicrobeWiki, su microbewiki.kenyon.edu.
- (EN, FR) Pasteur Institute, su pasteur.fr. URL consultato il 1º marzo 2009 (archiviato dall'url originale il 13 aprile 2007).